




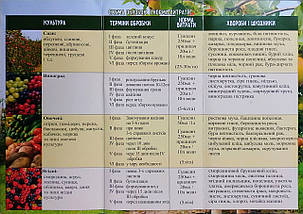



Гуапсин — вода для бактерій PCEUDOMONAS AUREOFACIENS
Інсексуальний Біопрепарат для боротьби з шкідниками та хворобами сільськогосподарських та інших культур, який містить у своєму складі штами PCEUDOMONAS AUREOFACIENS B-306 (IBM B-7096) і PCEUDOMONAS AUREOFACIENS В-111 (IM B-97), їхні метаболісти і дози макромементів (NP,K).
Препарат створений на основі реально існуючих в природі мікроорганізмів, який зменшує кількість фітофагов до безпечного рівня для всіх зелених насаджувань, на відміну від хімзлочинів, які знищують все живе. В результаті наукових досліджень, а також застосуванням ефективності Гуапсіна з грибковими захворюваннями 90-92%, з шкідниками 92-94%. Препарат нетокситний для людей, тварин, риб, бджіл, не накопичується в рослинах, ґрунті, не впливає на смак і колір вирощеної продукції.
Що ж до ліків, то бактерії потрапляють на рослину, а грибкові спори, які спричиняють хвороби або гриби, — патогенні.
У той же час бактерії Гуапсіна, які проникли в травну систему шкідників (поїдання листків або харчуючись соком), блокують їхню травну функцію, що зрештою призводить до загибелі.
Після обробки ґуапсином спостереження за рослинами виявили на стеблах і листях підвищену азотофіксову активність діазотрофових бактерій, які отримали назву асоціативної азотації. Рослини стимулюють діяльність бактерій за рахунок фотосинтез, що екскретизується (вироблені) у середовищі, ніж визначають солодку і сезонну динаміку азоту. Внося засоби - ми, по суті, створюємо штучний симбіоз.
Гаупсин сумісний з багатьма хімічними препаратами, у тому числі з гербіцидами (окрім меду та ртуть), застосування ліків на рослинах можна спостерігати за 24-36 годинами.
Результати досліджень
Вчені дійшли висновку, що бактерія PCEUDOMONAS AUREOFACIENS В-306 входить до складу Гуапсина Завдяки модернізації з’єднання ґрунтів, що розвиваються, рослини починають отримувати додаткове харчування. За короткий час після оприски починає рости фізіологічна активність самої рослини, а її корені всмоктують поживні речовини інтенсивніше.
Це відбувається завдяки рослинному гормону-індолілуксу, який виділяють бактерії.
Також псевдомонади, які ростуть у цьому ґрунті, придушують ріст бактеріологічних мікроорганізмів у різосфері, а також сприяють росту рослин. Внаслідок цього рослина отримує таку кількість азоту та інших поживних речовин, що ризосферні бактерії подібні до мінеральних добрив. Як показали експерименти внесення п’ятил. Гуапсина на 1 га. порівнюється з азотостимуляцією з 150-200 кілограмів. Селитри або 60-8 кг. Карбамида.
РИЗОСФЕРНІ БАКТЕРІЇ РОДА PSEUDOMONAS, СПОСОБЛИЧНІ РОСТУ І РОЗВИТКУ РАТЕНЬ
Гідність рослин до фітопатогенів залежить від їхньої взаємодії між кореневою системою рослин і різноманітними мікроорганізмами. Спорудження кореневої системи з ґрунтом — це складна екологічна ніша, заселена корисними, шкідливими та нейтральними для рослин мікроорганізмами.
Активна секреція коренів різних речовин забезпечує поживними субстратами мікроорганізми, які утворюють міцні сполуки як у кореневій тканині, так і на кореневій поверхні (різоплані), так і в ґрунті, що безпосередньо оточує коріння (різосфере).
З огляду на це в різосфері та різоплані в значній кількості концентруються бактерії, бактерії, бактерії, гриби, водорості та нематоди, значно перевищуючи кількість цих самих організмів у ґрунті. Однією з характеристик ризосфери різних рослин, що відбиває її засідання мікроорганізмами, є кількісне співвідношення R / S (rhizosphera / soil). Часто його визначають для різних видів, родів та сімей, які живуть на різосфері. Величина R / S показує, у скільки разів кількість мікроорганізмів певної таксономічної групи в різосфері цієї рослини перевищує кількість цих мікроорганізмів у ґрунті. Для більшості бактерій величина R / S коливається від 2 до 25. У різосфері, на відміну від вільної від коріння ґрунту, домінують грамотні бактерії, а флюоресцькі бактерії роду Pseudomonas. Деякі штами бактерій Pseudomonas putida, P. fluorescens, P. aureofaciens (chlororaphis), P. corrugata і д. сприяють значному поліпшенню росту і розвитку рослин. Нині бактерії, що мають сукупність корисних властивостей рослин, вважаються PGPR (від Plant Growth-Promoting Rhizobacteria — різобактерії, що сприяють росту рослин).
Дослідження цієї перспективної для практичного використання групи різобактерій викликають велике зацікавлення, що викликане регулярним проведенням міжнародних симпозіумів з проблеми вивчення і практичного використання PGPR. Серед PGPR різних таксономічних груп виділяється широкий набір корисних для рослин властивостей ризоферні PGPR Pseudomonas, які є потенційними об'єктами агробіотехнології для розробки на основі їх біологічних засобів захисту від фітопатогенів, а також біоперепаратів, які стимулюють ріст і підвищення продуктивності рослин.
Механізми позитивного впливу PGPR PSEUDOMONAS на рослини
Усі дослідники до цих пір, що мають позитивний вплив на псевдомонад рослини, можна умовно розділити на два типи: 1) прямий або безпосередній стимуляцію росту рослин за рахунок синтезу різних метаболітів, корисних для рослин; 2) По - перше, стимуляція росту рослин за рахунок витіснення та придушення ґрунтових фітопатогенів або мікроорганізмів, що пригнічують ріст рослин. Перш за все, до першого типу можна віднести здатність PGPR Pseudomonas синтезувати регулятори росту рослин і поліпшити фосфорне харчування рослин. Крім того, деякі штами псевдомонад здатні до фіксації атмосферного азоту та індукції рослин стійкості до фітопатогенів.
Пряма стимуляція росту рослин
Синтез регуляторів росту рослин. У різосферних псевдомонад найбільш добре вивчена здатність до синтезу індолів-3-уксусної кислоти, яка, як відомо, стимулює розвиток кореневої системи рослин. Однак більшість штамів псевдомонад, які стимулюють ріст рослин, продуковують іуК у невеликих кількостях (3 -5 мкг/мл), тоді як псевдомонади, що гноблять ріст рослин, продуцують до 20 мкг/мл ІуК. Дослідження синтезу IuК у PGPR Pseudomonas показують можливість отримання генетично модифікованих штамів PGPR Pseudomonas, які здатні до підвищеного синтезу IuК [3]. Перенесення в PGPR Pseudomonas деяких Плазмід біодеградації нафталіну також сприяє збільшенню рівня синтезу ІУК у різосферних псевдомонад, що викликане наявністьм у складі цих плазмид гена, відповідального за синтезу нафталініокази (перового ферменту в дорозі окислення нафталіну), що вступає в процес біосинтезу ІУК. Слід зазначити, що, окрім ІУК бактерії роду Pseudomonas, можуть продукувати інші регулятори росту рослин, як, наприклад, гібереліноподібні речовини.
Покращення фосфорного харчування рослин. Фос фор присутній у ґрунті у вигляді органічних (відкладів рослинного, тварини та мікробного походження) і неорганічних або мінеральних сполук. З цього загального пула фосфорних сполук лише близько 5% доступні рослинам. Деякі мікроорганізми, особливо міко- ризові гриби і деякі ризотоктерії, можуть посилювати ріст фосфору в рослини. Бактерії можуть використовувати дві системи підвищення концентрації екзегованого фосфату: 1) за рахунок гідролізу органічних фосфатів під дією фосфатаз; 2) розчиняючи мінеральні фосфати за рахунок продукції кислот. Бактерії роду Pseudomonas здатні до ефективного розчину фосфорних сполук, що можна використовувати для поліпшення фосфорного харчування рослин
Фіксація атмосферного азоту.
На даний час відомо трохи азотознавців псевдомонад і немає суворих доказів того, що їхній процес азоту вносить якийсь важливий внесок у стимуляцію росту рослин.
Проте роль PGPR Pseudomonas у процесі асоціативної азотфіксації активно обговорюється, оскільки такі штами можуть безпосередньо стимулювати ріст рослин, коли не існує фітопатогенів.
Цікаво, що в холодних кліматичних зонах у різосфері рослин азотчасті псевдомонади домінують над представниками інших таксономічних груп азотознавців. Перевага псевдомонад виражається в їх холодостійності, тому що оптимальна температура для азоту 14-20С. Водночас, для процесу азотифікації інших асоціативних діазотрофів, як, наприклад, бактерії роду Azospirillum, оптимальною температурою є 25extuss. Незважаючи на обмежену інформацію про роль азотифікації у стимуляції рослин псевдомонадами і слабку інтенсивність цього процесу у псевдомонадис, можна зробити висновок, що різактерії Pseudomonas разом з іншими незалежними і асоціативними діазофазіями Aтроbusacter, таким чином, а також, що, як і Azoliblibillibella, є антидемами.
Індукція резистентності до фітопатогенів.
Виготовлення рослин авірулентними або несумісними формами фітопатогенів може індуціювати різкість рослин до захворювань. Аналогічний ефект досягається в процесі інокуляції коренів рослин деякими штамами PGPR Pseudomonas. Наприклад, обробка коренів бобів і гвоздики певними штамами PGPR P. putida підвищує стійкість рослин до гриба Fusarium solani.
Це призводить до збільшення лігнифікації кореневої тканини і підвищення вмісту фітоаллексинів у стеблах рослин.
Під час індукції резолюції рослин іноді важко зрозуміти, чи це викликано прямим або опосередкованим впливом бактерій на рослині. Наприклад, штам P. fluorescens CHAО продуцує на додаток до антибіотиків та сідофорів ціаністичний водень і ефективно пригнічує захворювання тютюну, яке називається грибом Thielaviopsis basicola.
Накопичення геотермального водню в коренях індуцує стійкість тютюну до гриба, який виліковували вище, але в цілому він є результатом дії не тільки ціанистого водню, але й опосередкованих факторів (синтез седерфорів та антибіотиків).
Запобігання росту рослин
Бактерії роду Pseudomonas є однією з найбільш досліджуваних груп бактерій-атагоністів грунтових фітопатогенів. На даний час виділили багато штамів різосферних псевдомонад, які придушують або сповільнюють ріст і розвиток грибів та бактерій. Література, присвячена цій справі, настільки широка, що вона не дозволяє розглянути всі аспекти проблеми в межах цього дослідження. Механізми антагоністичних взаємодій псевдомонади та фітопатогенів дуже різняться. Розгляньмо два з них, найбільш досліджених і важливих з точки зору практичного використання різобактерій Pseudomonas.
Продукція сидерфорів. Дуже добре відомо про здатність псевдомонад продуцувати жовто-зелені флуорестичні водорозробні пігменти, які називаються сідофорами, що виконують функцію зв'язку і транспорту заліза в клітини бактерій. Сидерофори флюоресцьких псевдомонад мають різну хімічну структуру і, як правило, мають високий зв’язок з тривалентною залозою, утворюючи з ним стабільні комплекси, недоступні для використання фітопатогенами. Фітопатогени продукують власні сидерерифори, але, на відміну від сидофорів псевдомонад, вони набагато повільніше пов'язуються з іонами заліза, і псевдомонади виграють у конкурентній боротьбі з фітопатогенами за такий важливий елемент, як залізо. Отже, зв’язування заліза сідофорами псевдомонад призводить до обмеження росту фітопатогенів і поліпшення росту рослин.
Важливу роль сідофорів у антагоністичних взаємовідносинах різосферних псевдомонад із грунтовими фітопатогенами і стимуляції росту рослин неодноразово доведено під час інокуляції рослин штамами, які продукують сідофори, і їх мутантами, дефектними з синтезу сидерофорів. При цьому було помічено не лише супресуючу дію сидофорів на фітопатогенах, але й стимулюююючу дію на рослини. Це також підтверджується в експериментах з очищеним сідофором одного з штамів P. putida — B10 псевдобактином, який стимулює ріст картоплі.
Слід зазначити, що антагонізм псевдомонад щодо фітопатогенів, спричинених конкуренцією за залізо, впливає лише на низький вміст заліза в ґрунті. Різко падає захисний ефект сідофорпродуктивних штамів у кислих ґрунтах, де розчинність заліза та його доступність для всіх мікроорганізмів зростають. Надлишок заліза також призводить до репресії синтезу сидерфорів. Щоб розв'язати цю проблему, можуть дерепресовані мутанти сідерофорпродуючі штами, здатні до конститутивного синтезу сідерофорів, який не закладений від концентрації заліза в ґрунті. Можливість отримання таких регуляторних мутантів продемонстрована на одному з штамів PGPR Pseudomonas sp. – M114.
Синтез антибіотиків. PGPR Pseudomonas може продукувати широкий спектр метаболітів, у тому числі [4]. Найбільш дослідженими антибіотиками, які відіграють важливу роль у сприянні хворобам рослин, є антибіотики групи феназінів, флороґлюцинів, піолютеорин, пірролін та ооміцин А (ріс. 3, 4). Щоб захистити рослини від антибіотиків безпосередньо, було показано у дослідженнях Л. Томашів з бактерізації насіння пшениці штамом P. fluorescens 2-79 і його дефектним на синтезу феназінового антибіотика (феназин-1-карбонової кислоти) мутантом. Захисту від одного з збудників кореневої гнилості давав тільки первісний штам антибіотиків, тоді як мутантний штам хоча й не повністю, але в значній мірі втратив цю здатність. Слід зазначити, що мутанти PGPR Pseudomonas, які мають підвищений синтез антибіотиків, не поліпшували захисту рослин від фітопатогенів. Нещодавно була ототожнена генетична система, яка контролює біосинтез феназин-1-карбонової кислоти у штама P. fluorescens 2-79. Визначено нуклеотидну послідовність фрагмента геномної ДНК штаму 2-79 розміром 8505 п.н., і в секвенованій ділянці лікалізовані три опери - phzABCDEFG, phzI і phzR з області біосинтезу феназіну-1-карбонової кислоти. Вивчено роль окремих структурних генів у біосинтезі феназин-1-карбонової кислоти в клітинах E. coli, що містять клоновані гени phz, а також у відповідних мутантах P. fluorescens. Запропонована принципова схема руху біосинтезу цього антибіотика [5].
PGPR PSEUDOMONAS як агенти захисту рослин від фітопатогенів
Щоб захистити рослини від фітопатогенів, потрібно приблизно 70 років. Фахівці, які займаються цією проблемою, часто називають біологічний захист рослин за допомогою інших біологічних контролю фітопатогенів [6]. На початку 80-х років інтерес до біологічного контролю різко зріс у зв'язку з можливими біоперепаратами та біотехнологією, конкурентними хімічними засобами захисту рослин.
Гідроенергетична система псевдомона і фітопатогенів є основними, але не єдиними механізмами, які сприяють росту рослин. Дуже часто біологічний контроль фітопатогенів різосферними псевдомонадами є наслідком комплексної дії різних механізмів, що включає в себе не тільки синтезу сідефорів та антибіотиків, але й інші менш досліджені способи впливу різобактерій Pseudomonas на грунтових патогенів. При цьому, можливо, імовірний вплив псевдомонад на фітопатогени, як за рахунок синтезу сидофорів, антибіотиків та інших вторинних метаболітів, так і за рахунок простого конкуренції PGPR Pseudomonas і фітопатогенів за джерела азотного і вуглецевого харчування.
Одним з таких прикладів є штам P. fluorescens CHAO. Біологічний контроль гриба Thielaviopsis basicola спостерігається в результаті сумного ефекту щонайменше трьох метаболітів, які утворюються штамом CHAО, а саме: сідофора піординового типу, антибіотика 2,4-диацетилфлорогалюцина і цианістого водню.
Майже у всіх випадках біологічний контроль псевдомонадовими грунтовими фітопатогенами являє собою загальний ефект комплексних механізмів. За різних умов (состав ґрунт, температура, рН) переважають різні механізми.
Наприклад, якщо синтез сидофорів має кислий рН або підвищену температуру, то псевдомонас придушує і основні механізми біологічного контролю можуть стати синтезу антибіотиків або конкурентів з фібратогенами за джерела їжі. Цілком імовірно, що не всі механізми, які сприяють біологічному контролю фітопатогенів у різосферних псевдомонад, виявилися й вивчені.
Біологічний контроль псевдомонадами фітопатогенів in viro є і досі одним з основних критеріїв відбору практично корисних штамів PGPR Pseudomonas. Проте скринінг штамів-антагоністів in viro має істотний недолік. Антгонізм in viro і in vivo не завжди корелює, що вимагає обов'язкової перевірки штаму-антагоніста на дослідах на рослинах.
Таким чином, вже в даний час можливо ефективне використання штамів PGPR Pseudomonas, правильно підібраних до конкретних умов певного господарства в якості біологічних засобів захисту рослин, які є доповненням, а іноді і альтернативою хімічних засобів. Активно проведені дослідження в цьому напрямку і розроблені нові технології суттєво підвищать ефективність цих біоперепаратів.
| Основні | |
|---|---|
| Виробник | IBM |
- Ціна: 90 ₴
- Готово до відправки
- Код: Guap
Гуапсин Біопрепарат інсектофунгіцидний 1 літр
90 ₴
- +380 (99) 172-22-66
 Гуапсин Біопрепарат інсектофунгіцидний 1 літр
Гуапсин Біопрепарат інсектофунгіцидний 1 літр